

Affects of Viral Disease in Shrimp Farming
In Shrimp Disease control, they are Six viruses were known to affect Penaeid shrimp, but there are more than 20 viruses were identified as having affected wild stocks and commercial production. The OIE now lists seven viral diseases of shrimp in the Aquatic Animal Health Code, which are considered to be transmissible and of significant socio-economic and/or public health importance.
- These viral diseases as follows:
- White spot disease (WSSV).
- Yellowhead disease (YHV),
- Taura syndrome virus (TSV),
- Spawner-isolated mortality virus disease (SMV),
- Tetrahedral baculovirosis (Baculovirus penaei - BP),
- Spherical baculovirosis (Penaeus monodon-type baculovirus) and
- Infectious hypodermal and haematopoietic necrosis (IHHNV)
Penaeus vannamei and P. stylirostris are known to be carriers of the following viral diseases: WSSV, BP, IHHNV, REO, LOVV and TSV. These viruses can be transmitted to native wild Penaeid shrimp populations. Penaeus monodon are known carriers of: WSSV, YHV, MBV, IHHNV, BMNV, GAV, LPV, LOVV, MOV and REO.
Perhaps the biggest concern to Asian countries already or currently wanting to import P. vannamei is the possibility of introducing TSV. Despite original work suggesting Taura syndrome (TS) was caused by a toxic pesticide, it is now known that a single or perhaps several very closely related strains (mutations) of the Taura syndrome virus (TSV) are responsible for the TS pandemic in the Americas. TSV is a single strand RNA virus and hence susceptible to mutations, causing more concern, and is closely related to other insect viruses.
Taura Syndrome Virus was first identified from farms around the Taura river in Ecuador and subsequently spread rapidly to the whole of Latin and North America. TSV spread first throughout Ecuador and to Peru, Colombia (Pacific and Atlantic coasts), Honduras, Guatemala, El Salvador, Nicaragua, Hawaii, Florida and Brazil, Mexico, Texas, South Carolina and Belize and subsequently Asia including Mainland China and Taiwan Province of China and most recently Thailand probably through the regional and international transfer of live PL and broodstock P. vannamei.
Taura syndrome caused serious losses in revenue throughout Latin America. It has been suggested that TSV caused direct losses (due to shrimp mortality) and indirect losses due to loss of sales, increased seed cost and restrictions on regional trade were probably much higher. Taura syndrome so far appears to occur largely as a sub-clinical infection in populations of wild shrimp. Although P. monodon and P.japonicus appear largely unaffected, the potential impact of TSV on native stocks of P. indicus and P. merguiensis in Asia remains unknown, but a definite cause for concern.
The mechanism of spread of TSV is still uncertain, although initial theories concentrated on the spread through contaminated PL and broodstock between farms. Limited data have shown that TSV was introduced to Colombia and Brazil through contaminated broodstock from Hawaii. These broodstock were untested for TSV since it was not yet known that Taura syndrome had a viral cause. Such cases demonstrate once again more of the problems involved with transboundary movements of animals, even supposedly SPF ones. Recent research has shown that mechanical transfer through insect and avian vectors may be an equal or even more likely route of infection. TSV has sometimes been found in tissue bioassays of the water boatman (Trichocorixa reticulata), an estuarine insect common worldwide, and virus-containing extracts of this insect have been shown to induce infection in SPF P. vannamei under laboratory conditions. Patterns of the spread and mortality of P. vannamei in Texas have also suggested that the ingestion of infected insects is the probable mechanism of spread of TSV.

Infective TSV has also been demonstrated in the faeces of shrimp-eating seagulls (Larus atricilla) collected near ponds infected with TSV in Texas, USA. Experimental results have also shown that healthy shrimp can be infected through injection of cell-free homogenates prepared from infected shrimp, and by direct feeding on infected shrimp. Taura syndrome virus has also been shown to remain infective after one or more freeze-thaw cycles, indicating the possibility of regional transmission through infected frozen shrimp. With proper disinfection procedures and controls, however, this route is currently considered to be low-risk.
Taura syndrome virus is highly infective for P. vannamei, P. setiferus and P. schmitti. Penaeus stylirostris can be infected by injection, but appear to be highly refractory to TSV and have demonstrated tolerance to TS in growing areas affected by this disease. Other species including P. aztecus, P. duorarum, P.monodon, P. japonicus and P. chinensis have been experimentally infected, developed the disease and remained carriers, but show some resistance. Interestingly, like P. stylirostris, P. monodon and P. japonicas appear highly refractory to TSV, and although it retards growth rates, they remain asymptomatic and the virus has not yet been demonstrated to cause mortality in these species. However, since TSV is an RNA virus, with a high propensity to mutate, there is no guarantee that it will not mutate into a more virulent form for native Asian shrimp (as it did in Central America)
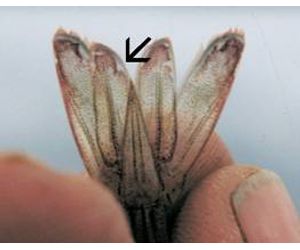
Taura Syndrome Virus has already been detected in P. vannamei in Mainland China and Taiwan Province of China with 19 cases reported to OIE from Taiwan Province of China in 1999, ten (resulting in 700 000 cases and 200 000 deaths) and seven (resulting in 500 000 cases and 50 000 deaths). Recently, TSV has been identified in Thailand. The Taura syndrome virus tends to infect juvenile shrimp within two to four weeks of stocking ponds or tanks (0.1-1.5g body weight) and occur largely within the period of a single moult cycle. In the acute phase of the disease, during pre-moult the shrimp are weak, soft-shelled, have empty digestive tracts and diffuse expansion of the red chromatophores, particularly in the tail (hence the common name - red tail disease). Such animals will usually die during moulting (5-95 percent), although the reasons for the large variability in survival rates remains unknown; adult shrimp are known to be more resistant than juveniles. Those shrimp that survive will show signs of recovery and enter the chronic phase of the disease. Such shrimp will show multiple, randomly distributed, irregular, pitted, melanised lesions of the cuticle. These gross or microscopic lesions will persist, but may be lost during moulting, the shrimp thereafter appearing and behaving normally. However, although the shrimp may then be resistant to recurrence of the disease, they often remain chronic, asymptomatic carriers of TSV for life, as has been shown by bioassays.
Standard histological and molecular methods may be used for detection, diagnosis and surveillance, although specific DNA probes applied to in situ hybridization assays with paraffin sections currently provide the greatest diagnostic certainty of this virus (OIE website). RT -PCR assays can also be used providing advantages of larger sample sizes and non-lethal sampling for broodstock. Additionally, live shrimp bioassays and serological methods with monoclonal antibodies can also be used for diagnosing infections with TSV. Eradication methods for TSV in culture facilities are possible and depend upon total destruction of infected stocks, disinfection of the culture facility, avoidance of reintroduction of the virus (from nearby facilities, wild shrimp and carriers) and restocking with TSV-free PL produced from TSV-free broodstock. Other methods suggested for controlling the virus include: switching to the refractory P. stylirostris, and (similar to those suggested for other viruses): maintenance of optimal environmental conditions, weekly applications of hydrated lime (CaOH) at 50 kg/ha, polyculture with fish (to consume dying and dead carriers) and development of TSV resistant lines of P. vannamei. In the past few years, considerable success has been achieved in the development of families and lines of P. vannamei which are resistant to TSV.
Most of the SPF P. vannamei suppliers from Hawaii and Florida now offer stocks of P. vannamei which have demonstrated resistance to TSV (SPF and SPR). Genetic selection programmes run throughout the Americas have also resulted in the production of SPR lines for TSV. The use of such SPR lines enabled the Latin American industry to recuperate from the worst of the TSV pandemic within three to four years. However, importation of such lines must be done with caution, since non-SPF animals, even though resistant to TSV, may still act as carriers and can result in the introduction of TSV into areas of Asia currently free from the disease. Aquacultural establishments, zones within countries, or countries that are considered TSV-free, are those which have been tested in an official crustacean health surveillance scheme for a minimum two years using the procedures without detection of TSV in any susceptible host species of shrimp 19. Additionally for aqua cultural establishments, they must be supplied with water that has been suitably disinfected and have barriers preventing contamination of the establishment and its water supply. New or disinfected facilities, may be declared free from TSV in under two years if all other requirements are met.
Whilst this degree of control may be possible in large-scale highly organized shrimp farms, the reality is that most farms are too small or disorganized to undertake such comprehensive measures. The lack of supporting infrastructure in regulation, testing and diagnosis is an additional constraint. This problem is not confined to Asia where farms are typically very small, but also occurs in Latin America where farms are far larger.
