

Affects of White Spot Syndrome Virus (WSSV) in Shrimp Farming (Part 3)
White Spot Syndrome Virus (WSSV) is now and has for some time been the most serious threat facing the shrimp farming industry in Asia. It is an extremely virulent pathogen with a large number of host species.
This disease is probably the major cause of direct losses of shrimp farming in Asia. Similarly, in Latin America, losses due to WSSV have been substantial. In addition, indirect losses in hatchery, feed and packing plant capacities and so on resulted in lost earnings. Similar problems have occurred throughout Central and South America, with the exception of Brazil and Venezuela, which remain WSSV-free due to the prompt and effective closure of their borders to all crustacean imports.The United States also managed to eradicate WSSV from its shrimp culture industry after initial losses through implementation of biosecurity measures, including the use of all SPF broodstock, although there are reports of its recent re-emergence in Hawaii.
WSSV is a large double-stranded DNA baculovirus. Other names for probably the same viral complex include Chinese baculovirus (CBV), White spot syndrome baculovirus complex (WSBV), Mainland China’s Hypodermal Hepatopoietic Necrosis Baculovirus (HHNBV), Shrimp Explosive Epidermic Disease (SEED), Penaeid Rod-shaped DNA Virus (PRDV), Japan’s Rod-shaped Nuclear Virus (RV-PJ) of P. japonicus, Thailand’s Systemic Ectodermal and Mesodermal.
Baculovirus (SEMBV) of P. monodon, red disease and white spot virus or disease. WSSV was first reported in farmed P. japonicus from Japan, but was thought to have been imported with live infected PL from Mainland China. At roughly the same time, it was discovered in cultured P. monodon, P. japonicus and P. penicillatus in Taiwan Province of China and then in P. monodon in southern Thailand. WSSV then spread rapidly throughout most of the shrimp growing regions of Asia, probably through infected broodstock and PL P. monodon. Then, in 1995, it was detected for the first time in farmed P. setiferus in Texas. It was also shown to be infective experimentally to both P. vannamei and P. stylirostris. WSSV did not reach the Philippines, which had an effective government ban on live imports, until an illegal introduction of Chinese PL P. monodon.
Other susceptible host species include the shrimp species P. merguiensis, Metapenaeus ensis, Metapenaeus monoceros and various crab species, whilst Palaemon setiferus, Euphausia superba, Metapenaeus dobsoni, Parapenaeopsis stylifera, Solenocera indica, Squilla mantis, Macrobrachium rosenbergii and a range of crab species can act as latent carriers, although Artemia appear unsusceptible. Later, in 1999, WSSV began affecting Latin America from Honduras, Guatemala, Nicaragua and Panama in Central America to Ecuador and Peru in the south and later to Mexico. The only shrimp farming countries to remain free of WSSV in Latin America are Brazil and Venezuela, who (like the Philippines) both placed immediate and effective bans on the importation of live crustaceans and developed their domestication programmes for producing virus-free seedstock.

The mode of transmission of WSSV around Asia was believed to be through exports of live PL and broodstock. The outbreaks in Texas and then Honduras followed by Spain and Australia, were thought to be due to the virus escaping from processing plants which were importing and processing frozen shrimp from infected parts of Asia, although this has never been proven. Regardless of their origin, isolates of WSSV have shown little genetic or biological variation, suggesting that the virus emerged and was spread from a single source. WSSV, as with most viral diseases, is not thought to be truly vertically transmitted, because disinfection of water supplies and the washing and/or disinfection of the eggs and nauplius is successful in preventing its transmission from positive broodstock to their larvae. Instead, it is generally believed that the virus sticks to the outside of the egg, since, if it gains entry to the egg, it is rendered infertile and will not hatch. Thus, using proper testing and disinfection protocols, vertical transmission can be prevented in the hatchery, as proven by the Japanese who to date have successfully eliminated WSSV from captive stocks in the country through disinfection and PCR checking of broodstock and nauplii)
Using mathematical epidemiology modelling, Soto and Lotz (2001) showed that WSSV was more easily transmitted through ingestion of infected tissues than through cohabitation with infected hosts, and that P. setiferus was much more susceptible than P. vannamei to infection. Although it is clear that live Penaeids can carry the virus and infect new hosts through reproduction (transmission from broodstock to larvae), consumption or cohabitation with diseased or latent carriers, and that it is possible for frozen shrimp to be infective, other modes of transmission are also possible. For example, Australia is considered WSSV (and YHV)-free, although WSSV was detected in the Northern Territories in 2000 associated with imported bait shrimp, before being eradicated.
Data regarding the presence and effects of WSSV in wild shrimp populations in infected countries is scarce, but it is known to be present in wild shrimp in both Asia and Latin America. WSSV infects many types of ectodermal and mesodermal tissues, including the cuticular epithelium, connective, nervous, muscle, lymphoid and haematopoietic tissues. The virus also severely damages the stomach, gills, antennal gland, heart, and eyes. During later stages of infection, these organs are destroyed and many cells are lysed. The shrimp then show reddish colouration of the hepatopancreas and the characteristic 1-2mm diameter white spots (inclusions) on their carapace, appendages and inside surfaces of the body. They also show lethargic behavior and cumulative mortality typically reaches 100 percent within two to seven days of infection.
Increasingly, since the late 1990s, it has become clear that the presence of WSSV in a pond does not always lead to disaster. Work in Thailand has shown that outbreaks are usually triggered from latent P. monodon carriers by some environmental changes, probably related to osmotic stress through changes in salinity or hardness or rapid temperature changes. Similarly in Latin and North America, fluctuations in temperature have been shown to induce mortalities of infected P. vannamei. However, there have been conflicting reports about constant temperatures which have been reported to: limit mortality due to WSSV at 18 ºC or 22 oC and induce 100 percent mortality at 32 oC in the US, yet induce mortality at less than 30 oC and protect from it at greater than 30 oC in Ecuador
Additionally, three to four years of genetic selection work (selection of shrimp surviving WSSV outbreaks) on the domesticated stocks of P. vannamei appear to have resulted in enhanced resistance to WSSV in Ecuador. Thus the culture industries for P. vannamei in Central and South America have been slowly recuperating since the start of the WSSV epidemic in 1999. For example, Ecuador was exporting 115 000 metric tonnes in 1998, which dropped to only 38 000 metric tonnes in 2000 after the arrival of WSSV in 1999. Subsequently, Ecuador has recovered to export an estimated 50000 metric tonnes in 2003.
Prevention methods are similar to those with TSV. All live and frozen shrimp should be checked by PCR prior to importation from infected areas to those currently disease-free. Broodstock should be PCR screened before breeding. PL should also be PCR screened before stocking into ponds, as this has been proven to result in a higher percentage of good harvests. PCR is not an infallible method for detection of WSSV, but it is the best diagnostic procedure currently available. Washing and disinfection of eggs and nauplii have also been shown to prevent vertical transmission of WSSV from infected broodstock to larval stages. Feeding with fresh crab and other crustaceans to broodstock should be avoided. Polyculture techniques with mildly carnivorous fish species (such as Tilapia spp.) have also proven effective at limiting the virulence of WSSV in ponds, as the fish can eat infected carriers before they become available to the live shrimp.
The white spot virus only remains viable in water for 3-4 days, so disinfection of water used for changes and fine screening is effective in preventing transmission. Dose rates of 70ppm formalin have been shown to prevent transmission and not cause any harm to shrimp. In addition, all effluent from farming or processing operations with the possibility of WSSV infections should be disinfected (i.e. with formalin or chlorine) prior to discharge.
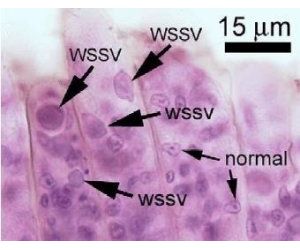
WSSV can be detected by using PCR, or with probes for dot-blot and in situ hybridisation tests. It can also be visually diagnosed through the presence of the characteristic white spots (although these are not always present in infected animals). WSSV can be confirmed histologically (particularly for asymptomatic carriers) by the presence of large numbers of Cowdrey A-type nuclear inclusions and hypertrophied nuclei in H&E-stained sectioned tissues, or simply by rapid fixation and staining of gill tissue and microscopic examination.